






Cytochemists study membranes either by first splitting them along the plane of the two lipid leaflets, inbetween the lipid bilayer. Or they may label transmembrane proteins and watch the dynamics of binding, membrane fluidity, or membrane uptake. Another favorite approach to membrane function is a study of specialized junctions formed at key membrane sites.
How do you prove membranes are fluid?
The first group of studies in this presentation will focus on membrane fluidity. This is made possible by the rotation of lipids and proteins in the bilayer. Your text reading for this section is found on pages 498-501 in Alberts et al. Molecular Biology of the Cell, Third Edition, Garland Publishing, N.Y., 1994

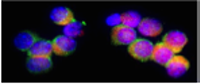
Membrane proteins (like lipids) rotate about an axis perpendicular to the plane of the bilayer. However, they may also move laterally. The following experiments show that not only do membrane proteins move, they can rearrange. The first experiment was done on fused cells. One group of cells came from a mouse line and the other from a human cell line. The investigators obtained antibodies to proteins in the membrane and conjugated these antibodies to fluorescein (green) or rhodamine (red) fluorescent compounds. The cells were then fused, forming a "heterocaryon" (note the two nuclei). Then, the two sets of antibodies were applied. Initially the cells were labeled half red and half green. However, after 40 min at 37 C, the labels mixed and the cells showed mixing of the two dyes. Often this mixing turns out to be white fluorescence.
This figure was taken from Alberts et al. Molecular Biology of the Cell, Third Edition, Garland Publishing, N.Y., 1994, Figure 10-34.
Note: if incubation was at 4 C, what do you think might happen?
Photobleaching experiments
In the next experiment, the label on the membrane is "bleached" in one area with a laser beam. The figures to the right show the results. Bleaching removes the label on the protein and results in a clear area (area of no label) in a completely labeled cell (see following figure).
Then, one can watch the recovery in the bleached area as the proteins + label diffuse into the irradiated patch. The figures show the bleached area as white. In the figure below, the recovered patch is a lighter red.


One can measure recovery in the bleached region with time after bleaching. The fluorescence decreases dramatically right after laser bleaching. Then, there is a gradual recovery with time.
The figures were taken from Alberts et al. Molecular Biology of the Cell, Third Edition, Garland Publishing, N.Y., 1994, Figure 10-36
Domains and Microdomains: How membranes support cell polarity, signaling, and unique functional sites.
Polarized cells have one side facing a special region in an organ and the other side facing another region. For example, in the intestine, the lining cells are polarized in that they express distinct structural and functional domains. See this site for more detailed information. The side responsible for absorption of nutrients faces the lumen. The lateral surfaces are specialized with barriers to entry and the basal surface is specialized to send nutrients and water to the blood stream. Each specialization is conferred by proteins associated with the membrane.
This figure below was taken from Alberts et al. Molecular Biology of the Cell, Third Edition, Garland Publishing, N.Y., 1994, Figure 10-37.
One can prevent mobility of proteins in a lipid bilayer either by structures in or on
the cell, or by experimental means. For example, as we will see in the section on tight junctions, apical proteins may be tethered
and prevented from entering the lateral or basal domains. The tight junction that actually
fuses the membranes  in places may help to prevent entry as well, although this may
be controversial.
in places may help to prevent entry as well, although this may
be controversial.
Return to Menu
Earlier in this presentation, we discussed the interaction of sphingomyelins and cholesterol forming rafts or platforms that allow protein attachment. This helps organize the membrane into functional microdomains. A microdomain can be considered a functional unit that involves different types of proteins, enzymes etc. Certain types of junctions or signaling cascades could work out of microdomains. The above figure from your text shows ways that membranes can be organized into domains and microdomains.
As shown in the first text figure, above: complexes of proteins such as receptors, those involved in second messenger signaling, or those in specialized junctions are often aggregated in one region of the cell perhaps by interactions with sphingomyelins and cholesterol platforms or rafts.

In the middle figures:
- Proteins can be tethered by interactions with macromolecules outside the cell (like extracellular matrix), or inside the cell (for example, cytoskeletal filaments).
- In the right hand figure, a complex of proteins can interact with proteins on another cell and be tethered in this way.
This figure was taken from Alberts et al. Molecular Biology of the Cell, Third Edition, Garland Publishing, N.Y., 1994, Figure 10-39
An example of such an interaction can be found in one type of junction, called the gap junction.
Gap Junctions
This type of junction allows communication between cells. This type is called a gap junction. Small molecules or ions can pass through, as we will see by the following figures.

The above freeze-fracture /freeze etch image shows the internal view of the gap
junction on the left. The proteins look like little donuts which reflects the fact
that they are actually a channel. These proteins are "connexon" molecules. The
side facing the cytoplasm (called the P face) is shown in the center panel. The region
looks like aggregated lumps. Finally, the typical electron microscopic view is seen in the
third panel. This shows a thin line between the two plasma membranes indicating a
"gap junction".
This figure was modified from Bloom and Fawcett, A Textbook of Histology, Chapman and Hall, N.Y., Twelfth Edition, 1994, Figure 2-14.
Gap junctions are formed by the dimerization of 6 connexin molecules to form a
hemichannel called a "connexon". These are brought to the cell surface as a unit
and one hemichannel docks with another hemichannel on an adjacent cell. Alignment of
the pore regions allow transport of small molecules from cell to cell. 
There are several ways to prove the cells are communicating by gap junctions. First, one can identify the connexon molecules by immunocytochemical labeling. Second, one can identify the actual junctional complex with freeze-fracture/freeze etch. To see if they are functional, however, one needs to inject one cell with a dye and watch to see if it is transferred to another cell. This cartoon diagrams a view of a gap junction showing molecules that can freely pass. Ions pass and in this way the cells can be electrically coupled together. Other small molecules that pass through include cyclic AMP (a second messenger) and the dye marker fluorescein. This last compound enables the scientist to study transport through the gap junction.
This figure was modified from Bloom and Fawcett, A Textbook of Histology, Chapman and Hall, N.Y., Twelfth Edition, 1994, Figure 2-15.
To learn how modern Cell Biologists study connexins and the formation of gap junctions, review Jordan, K et al. "Trafficking, Assembly and Function of a Connexin43 Green Fluorescent Protein Chimera in Liver mammalian cells." Molecular Biology of the Cell. 10: 2033-2050, June, 1999.
Abstract:
To examine the trafficking, assembly, and turnover of connexin43 (Cx43) in living cells, we used an enhanced red-shifted mutant of green fluorescent protein (GFP) to construct a Cx43-GFP chimera. When cDNA encoding Cx43-GFP was transfected into communication-competent normal rat kidney cells, Cx43-negative Madin-Darby canine kidney (MDCK) cells, or communication-deficient Neuro2A or HeLa cells, the fusion protein of predicted length was expressed, transported,and assembled into gap junctions that exhibited the classical pentalaminar profile. Dye transfer studies showed that Cx43-GFP formed functional gap junction channels when transfected into otherwise communication-deficient HeLa or Neuro2A cells. Live imaging of Cx43-GFP in MDCK cells revealed that many gap junction plaques remained relatively immobile, whereas others coalesced laterally within the plasma membrane. Time-lapse imaging of live MDCK cells also revealed that Cx43-GFP was transported via highly mobile transport intermediates that could be divided into two size classes of <0.5 um and 0.5-1.5 um. In some cases, the largerintracellular Cx43-GFP transport intermediates were observed to form from the internalization of gap junctions, whereas the smaller transport intermediates may represent other routes of trafficking to or from the plasma membrane. The localization of Cx43-GFP in two transport compartments suggests that the dynamic formation and turnover of connexins may involve at least two distinct pathways.
Return to Menu
Detection of Membrane Receptors
Receptors are a special class of transmembrane protein that
can be detected by their ligand. They have the reactive group
projecting from the surface that binds a specific ligand. This cartoon shows the
interaction between the ligand secreted by one cell (such as a hormone) and its receptor.
The lower cartoon shows interactions between two cells, one with a receptor for a protein
on the surface of the other cell. This kind of reaction may occur when natural killer
cells recognize a foreign cell.

What do you expect to happen if the cell is cooled to 4 C?
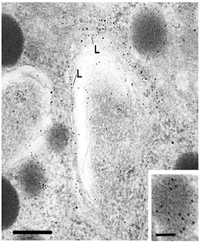
The figure to the right shows how a receptor-ligand complex can be detected at the light microscopic level. The ligand is attached to biotin and then it binds the target cell. It is detected by an avidin peroxidase conjugate which is visualized by a reaction for the enzyme peroxidase. This protocol is called "affinity cytochemistry". In this figure, the biotinylated ligand is labeled black (note that it is capping on one of the cells). The orange label defines the protein in the cell by immunocytochemistry. For more information, consult our Affinity Cytochemistry Web page

In a later lecture, we will learn the sequence of reactions that follow receptor binding. Usually the binding causes aggregation (patching) of the receptor-ligand complex. Then the aggregates accumulate at one pole (opposite to that of the centrioles) in a "cap" (called capping). At this point they may be internalized and either degraded or used for internal functions. All of this is possible because the membrane is fluid and allows lateral movement of theproteins. This figure was taken from Alberts et al. Molecular Biology of the Cell, Third Edition, Garland Publishing, N.Y., 1994, Figure 10-35
For more information, contact:
Gwen Childs, Ph.D.,FAAA
Professor and Chair
Department of Neurobiology and Developmental Sciences
University of Arkansas for Medical Sciences
Little Rock, AR 72205
For questions, contact this email address:
