
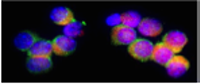








Intermediate filaments are important components of the cell's cytoskeletal system. They may stabilize organelles, like the nucleus, or they may be involved in specialized junctions. They are distinguished from "thin filaments" by their size (8-10 nm) and the fact that thin filaments are obviously motile. However recent evidence indicates that Intermediate Filaments may also have dynamic properties. See Side Bar for some photographs.
Intermediate filaments are one of three types of cytoskeletal elements. The other two are thin filaments (actin) and microtubules. Frequently the three components work together to enhance both structural integrity, cell shape, and cell and organelle motility. Intermediate filaments are stable, durable. They range in diameter from 8-10 nm (intermediate in size compared with thin filaments and microtubules). They are prominent in cells that withstand mechanical stress and are the most insoluble part of the cell. The intermediate filaments can be dissociated by urea.
There are five different types of Intermediate filaments:
- Types I and II: Acidic Keratin and Basic Keratin, respectively. Produced by different types of epithelial cells (bladder, skin, etc).
- Type III. Intermediate filaments are distributed in a number of cell types, including: Vimentin in fibroblasts, endothelial cells and leukocytes; desmin in muscle; glial fibrillary acidic factor in astrocytes and other types of glia, and peripherin in peripheral nerve fibers.
- Type IV Neurofilament H (heavy), M (medium) and L (low). Modifiers refer to the molecular weight of the NF proteins. Another type IV is "internexin" and some nonstandard IV's are found in lens fibers of the eye (filensin and phakinin).
- Type V are the lamins which have a nuclear signal sequence so they can form a filamentous support inside the inner nuclear membrane. Lamins are vital to the re-formation of the nuclear envelope after cell division.
Intermediate Filament Polymerization.
The monomer:
Each intermediate filament monomer consists of an alpha helical rod domain, which connects the amino (head) and carboxyl( tail) terminals. The figure below (16-12 from Alberts et al Biology of the cell, Garland Publishing, N.Y. 1996) shows some examples of monomers.

Formation of the protofilament

The rods coil around another filament like a rope to form a dimer. The N and C terminals of each filament are aligned. Some Intermediate filaments form homodimers; other form heterodimers.

These dimers then form staggered tetramers that line up head-tail. Note that the carboxy and amino terminals project from this protofilament. This tetramer is considered the basic subunit of the intermediate filament.

The final 10 nm filament is a helical array of these tetramers.
Regulation of Assembly or disassembly of intermediate filaments.
Most of the intermediate filaments are fully polymerized. However, there is evidence that even these stable structures have dynamic properties. There is some free tetramer in the cytoplasm as if this is the basic subunit for assembly of new filaments. Also, if one phosphorylates serine residues in the amino terminals one can cause disassembly.
Add labeled tetramer to a cell that produces that type of intermediate filament. Can watch the tetramer become incorporated into the cytoskeletal system and the label is seen in a linear or squiggle array. If you add it to a cell that does not normally produce the tetramer, then it will not add and the cytoskeletal system will not "light up".
This can be tested by Fluorescence recovery after photobleaching (FRAP)
This technique uses UV laser light to bleach an area of label in a cell. Then, one can time the recovery of the label either following introduction of new labeled material, or via simple movement of label into the structure. In the case of the Intermediate filaments, FRAP allows one to detect incorporation of labeled tetramers into a bleached spot in the cytoskeleton. One can compare times of incorporation of different types of tetramers into different types of Intermediate filaments. One can also observe motility of these structures. The paper that will be read for this lecture shows examples of both of these types of tests. In the cell below, a dark spot forms after laser photobleaching. The spot is smaller after 30 min and almost gone after 2 h.

Intermediate Filament Associated Proteins
Intermediate filament associated proteins may bind filaments in cross linked (to improve stability), or they may bind the filaments to other structures. Some examples are seen below.
- Plectin: Cross links with microtubules
- Lamin receptor B: binds to inner nuclear membrane
- Ankyryn: binds actin to Intermediate filaments at base of cell
- Desmoplakin: binds Intermediate filaments at site of desmosome
Types of Intermediate filaments
Lamins

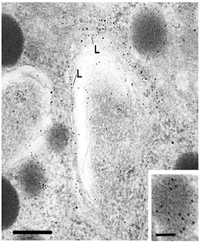
In evolution, Lamins were probably the first intermediate filaments made. They have a very long rod domain and carry a nuclear transport signal because they reside in the nucleus just under the nuclear envelope. They are continuous except for a break at the sites of the nuclear pore complex.

Above they are shown to form a lattice-like array (from Alberts et al, Garland Press, NY). The figure to the left is an electron micrograph of the area containing the lamins, just inside the nuclear envelope. They are difficult to distinguish from the nearby dense heterochromatin.
Lamins are phosphorylated at the end of prophase and this causes them to disassemble as the nuclear envelope also breaks down. Then, the phosphate is removed just before the nuclei of the daughter cell forms and the lamin filaments reassemble around each set of chomosomes, under each daughter cell's inner nuclear membrane. One can block this process by adding antibodies to lamins before the nuclear membranes are formed.
Specialized junctions
Types I and II intermediate filaments are acidic and basic keratins, respectively. Their monomers are found in the same cells and the dimers must contain one of each type (a heterodimer). If one gives labeled monomers of only one type, few cells will add the label to the cytoskeletal system. However if one gives monomers of both types, the keratin cytoskeletal systems will be heavily labeled.
Keratins also have subtypes that are unique to different epithelial cells (bladder, skin, etc) or even different subsets of one cell type (like basal epidermal cells). This is useful in detection of the origin of cells in a tumor, especially cells that have metastasized.
In epithelia, keratins intermediate filaments form junctions that hold cells together (desmosomes), or attach cells to matrix (hemidesmosomes). In muscle cells, the intermediate filaments that form the desmosome are "desmins".
Desmosomes: Two plaques on adjacent cells (containing desmoplakin and other proteins) are connected by cadherin molecules. These molecules are linked by calcium. The intermediate filaments loop into the plaques spreading out into the cytoplasm. This links two cells together structurally. Keratins



The above cells are from skin and the cells look as if they have projected spines that touch spines from adjacent cells. Actually these are sites of desmosome connection which is a vital junction in the skin. The fixation technique has caused the cells to shrink, leaving the connection sites visible. An electron micrograph showing a desmosome is seen to the left. The intermediate filaments are looping in in almost a parallel fashion.
Patients who make antibodies to cadherin molecules will have weak or absent desmosomes and the skin will form blisters. These fluid filled areas will lie in the regions where the cells with spines are found.
Hemidesmosomes: Are sites of connections at the base of an epithelial cell with the matrix. The cartoon below shows the components. Intermediate filaments are stuck in a plaque (like the desmosome plaque) and Integrin molecules (receptors for matrix proteins) help connect the site with the matrix.
 ‘
‘
Type III Intermediate Filaments
Found in a variety of cell types. Each is unique to that cell type and used to identify tissue containing that cell type. Vimentin is found in cells derived from mesoderm: fibroblasts, endothelial cells, white blood cells;
Desmin is found in muscle cells, connecting Z disks and may connect center of contractile units. It is also found connecting to desmosomes in specialized junctions (cardiac muscle).
Glial fibrillary acidic protein is found in glial cells in the central nervous system.
Type IV Intermediate filaments
Include Neurofilament L, M, or H (named for low, medium or high molecular weight. These neurofilaments are linked by plectin cross bridges to each other and to microtubules. This adds to strength and spacing.
Neurofilament proteins add to the diameter of the axon and therefore influence its function (larger axons conduct faster).
For more information, contact:
Gwen Childs, Ph.D.,FAAA
Professor and Chair
Department of Neurobiology and Developmental Sciences
University of Arkansas for Medical Sciences
Little Rock, AR 72205
For questions, contact this email address:
