







Tier I shows budding from ER that is arranged facing a central zone at one end of the Golgi complex. These buds become vesicles and are coated with COPII protein coats. The above drawing shows an actual interface between the ER and the Golgi complex. The "Export complex" is seen at the top of the drawing. Note that the vesicle are moving to contribute to the cis-Golgi network of vesicles and cisternae.

The movement of these special transport vesicles is an energy requiring process. If one blocks production of ATP, the transport will not happen. This drawing shows how the rough endoplasmic reticulum forms vesicles (without ribosomes attached) that carry the newly synthesized proteins to the Golgi complex.
The inside of the vesicle becomes continuous with the inside of the Golgi cisternae, so that protein groups pointing towards the inside, could eventually be directed to face the outside of the cell.
Carbohydrate groups are attached and any subunits may be joined in these cisternae. The protein is then passed to the final region of the Golgi called the "trans face". There it is placed in vacuoles that bud from this region of the Golgi complex. These may be a certain size or density, characteristic of the cell itself. The vacuoles continue to condense the proteins and the final mature secretory granule is then moved to the membrane for secretion.

This electron micrograph illustrates a Golgi Complex. It is curved with its Trans face pointing away from the nucleus toward the cell periphery. The numerous vesicles in the area are transporting the proteins to and from cisternae.

Transport of material in and out of the Golgi complex involves budding and fusion of vesicles. This cartoon shows that the membranes of each join and align themselves during the process so that the inside face remains in the lumen and the outside face remains towards the cytoplasm.
Return to Menu
What types of secretion are controlled by the Golgi Complex?
The Golgi complex controls trafficking of different types of proteins. Some are destined for secretion. Others are destined for the extracellular matrix. Finally, other proteins, such as lysosomal enzymes, may need to be sorted and sequestered from the remaining constituents because of their potential destructive effects. This figure shows the two types of secretory pathways. The regulated secretory pathway, as its name implies, is a pathway for proteins that requires a stimulus or trigger to elicit secretion. Some stimuli regulate synthesis of the protein as well as its release. The constitutive pathway allows for secretion of proteins that are needed outside the cell, like in the extracellular matrix. It does not require stimuli, although growth factors may enhance the process.

Finally, this cartoon also shows the packaging of lysosomes which will be discussed in more detail in a later section. For more information go to the lysosome web page.
Return to Menu

As you will recall from a previous presentation on rough endoplasmic reticulum we saw that they are inserted in the membrane at the level of the rough endoplasmic reticulum. The protein sequence is coded for membrane insert start and stop sites. This directed the insertion and alignment points. Those that are multipass proteins have multiple start and stop sites.
The important role of the Golgi Complex is to make certain the plasma membrane proteins reach their destination. This figure shows the route. Note that the orientation of the protein is maintained so that the region destined to project outside the cell (a receptor binding site, for example), ends up in that place. In order to do this, it must be placed so that it faces inside the vesicle. Return to Menu
How does the Golgi complex add carbohydrate groups to a glycoprotein?

The Golgi Complex is compartmentalized. Phosphorylation occurs in the Cis region. In other regions, different types of carbohydrates are added as a glycoprotein passes through the cisternae. This figure illustrates the different regions where sugars like mannose (man), galactose (gal), etc are added. The final sorting is done in the Trans Golgi complex.
The functional differentiation of the Golgi complex can be studied with the electron microscope with specific techniques that detect enzymes. The cis region is rich in lipid-bearing membranes and can be delineated by osmium tetroxide labeling. The middle regions label for enzymes that add carbohydrates or other groups on the product. The inner, or Trans region, is the area where the lysosomes are sorted. Therefore, it is heavily labeled for acid phosphatase.
There is much interest in understanding how the different Golgi cisternae are organized and differentiated. A number of models exist, however a favorite is called the "Maturational model" (Bannykh S.I. and Bakch, W.E. Membrane Dynamics at the Endoplasmic Reticulum Golgi Interface J Cell Biol 138: 1-4 1997)
This model suggests that the new vesicles from the ER enter the cis Golgi network and retrograde vesicles (bearing COPI) coats move to merge with the cis region cisternae. These carry Golgi complex processing enzymes and their targeting to this region may be dependent on the low concentration of these processing enzymes. Then, as processing continues, the middle cisternae contain more mature product and lower amounts of the enzymes needed in the beginning. Finally, the trans region is specialized for sorting, containing receptors to sort and isolate lysosomal enzymes, for example.
For more information about modern studies that address this question, see: Wooding, S and H.R.B. Pelham, The dynamics of Golgi protein traffic visualized in living yeast cells. Molecular biology of the cell. 9: 2667-2680 1998

Sometimes vital proteins needed in the rough endoplasmic reticulum are transported along with the other proteins in the Golgi complex. The Golgi complex has a mechanism for trapping them and sending them back to the rough endoplasmic reticulum.

This cartoon shows the process. The protein destined for secretion is red. The blue protein must remain in the rough endoplasmic reticulum. The rough endoplasmic reticulum has inserted a receptor protein on the membrane it sends to the Golgi complex in the transitional vesicles (shown in green). These are retrograde vesicles and are therefore coated with "COPI" (coatamer). The ER protein receptor captures all of the protein that carries the ER residency signal. . Vesicles then bud from the Golgi complex and move back to the rough endoplasmic reticulum. The receptor can circulate and continue to return the proteins needed by the endoplasmic reticulum.
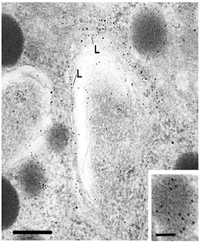
A drug called "brefeldin A" blocks the transfer of protein to the Golgi complex, however the reverse transport is not blocked. The following electron micrograph shows the results of an experiment after a brefeldin A block.
For more information, see also, Cole, N.B., Ellenberg, J, Song, J, DiEuliis, D and Lippincott-Schwartz, J. Retrograde transport of Golgi-localized proteins to the ER. J Cell Biol 1-15, 1998.

For more information, contact:
Gwen Childs, Ph.D.,FAAA
Professor and Chair
Department of Neurobiology and Developmental Sciences
University of Arkansas for Medical Sciences
Little Rock, AR 72205
For questions, contact this email address:
